

Hannah Warlick1*
1 Department of Psychology and Neuroscience, The University of North Carolina at Chapel Hill, 225 E. Cameron Avenue, Chapel Hill, NC 27599-3270
* hwarlick72803@gmail.com
Abstract
Adolescence is a critical period for basal ganglia maturation marked by increased vulnerability to depression. Using a previously collected longitudinal magnetic resonance imaging (MRI) and self-report dataset from 167 adolescents (age 11.9-18.6), we analyzed information across four timepoints spanning five years. We examined associations between depressive symptoms and age-related changes in tissue iron, an indirect marker of dopaminergic function, using multi-level modeling. Tissue iron levels increased with age across basal ganglia regions (caudate, putamen, nucleus accumbens and pallidum), with a stronger age-related effect in the caudate among females. Within-person changes in depressive symptoms had no significant effect on tissue iron trajectories. However, adolescents with higher average depressive symptom severity demonstrated greater developmental increases in caudate tissue iron. This unexpected pattern suggests that elevated depressive symptoms may coincide with accelerated maturation of caudate dopaminergic systems, reflecting influences such as early life stress and pubertal timing. These findings indicate that developmental patterns of basal ganglia iron vary with depressive symptom severity and sex during adolescence. The results indicate the importance of individual differences in reward-related neurodevelopment and identify the caudate as a potential biomarker target for future studies examining the neural correlates of adolescent depression.
Introduction
Adolescence is a critical period of brain development, marked by the maturation of subcortical structures, including the basal ganglia (Figure 1; Mills et al. 2021; Wierenga et al. 2014), which play a significant role in reward processing and motivation (Galvan 2010; Yoshida et al. 2022). During adolescence, basal dopamine release decreases while dopamine storage capacity increases, allowing the larger dopamine responses to rewarding stimuli (Galvan 2010). Researchers believe this reallocation of dopamine enhances reward salience, sensitivity and risk-taking behavior during adolescence (Urošević et al. 2012; Wahlstrom et al. 2010).
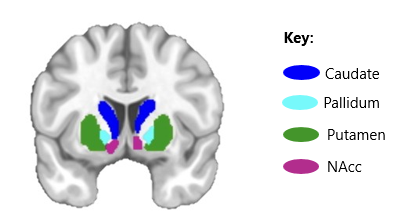
Figure 1. Key structures of the basal ganglia.
Iron plays a critical role in this process, serving as an essential cofactor in dopamine synthesis (Daubner et al. 2011). The basal ganglia, particularly the putamen and pallidum, are rich in iron, which accumulates across adolescence (Hill and Switzer 1984; Larsen et al. 2020a). This increase may support the expansion of dopamine storage capacity and enhances the dopaminergic responses (Galvan 2010; Laviola et al. 2003). Examination of tissue iron offers insight into adolescent dopamine-related function.
The basal ganglia is responsible for mood regulation via reward-processing circuits.. Adolescents with major depressive disorder exhibit suppressed reward responses and altered functional connectivity between the striatum and midline cortical structures (Davey et al. 2008; Gabbay et al. 2013; Luking et al. 2016; Rappaport et al. 2020). Depression is one of the most prevalent forms of psychopathology in adolescents, with approximately 34% of individuals aged 10 to 19 reporting clinically significant symptoms (Shorey et al. 2022). Anhedonia, or reduced pleasure in response to rewards, is a core symptom of depression and has been linked to dysfunction in reward-processing neurocircuitry (Cooper et al. 2018; Gabbay et al. 2013; Rappaport et al. 2020). Reduced activity in the nucleus accumbens and altered striatal connectivity are associated with increased depression severity and anhedonia (Gabbay et al. 2013; Rappaport et al. 2020).
Iron deficiency has been linked to depressive symptoms, with lower brain and peripheral iron levels associated with higher depression severity across the lifespan (Shah et al. 2021; Stewart and Hirani 2012; Yi et al. 2011; Vahdat Shariatpanaahi et al. 2007; Wu et al. 2023; Yao et al. 2017). These findings suggest that individual differences in tissue iron may relate to adolescent depression severity via effects on dopaminergic function.
To date, no studies have examined longitudinal associations between depressive symptoms and developmental trajectories of basal ganglia tissue iron across the duration of adolescence. Understanding this relationship is critical, as depression emerging during this period can have long-term effects on academic, social and occupational outcomes (Modrzejewska et al. 2022). Further knowledge of adolescent brain development could inform interventions to reduce long-term negative impacts of depression. This study investigates associations between depressive symptoms and tissue iron development across adolescence. We aim to replicate age-related increases in basal ganglia tissue iron and determine whether individual differences in these trajectories relate to depressive symptom severity. We hypothesize that adolescents with smaller age-related increases in tissue iron will exhibit greater depressive symptoms, whereas those with larger increases will show fewer.
Methods
Participants
The dataset and procedures for recruitment, data collection and processing used in this study were originally reported in Flannery et al. (2025). Participants were middle school students (6th and 7th grade) at the time of enrollment from a small but demographically diverse rural community in North Carolina. Data were collected between December 2016 and January 2022. Adolescents were excluded from the functional magnetic resonance imaging (fMRI) scan for braces or a permanent top retainer, claustrophobia, metal in the body, certain learning disabilities or a history of seizures or head trauma. Those prescribed medication for attention deficit/hyperactivity type disorders were asked to not consume their medication 24 hours prior to scanning. Participants with missing fMRI or self-report data across all timepoints were excluded from further analysis. The final sample included 167 adolescents (51.5% female; ages 11.9-18.6 years; 6th-11th grade) with 468 fMRI timepoints across four waves of data collection over five years. Appointments for data collection took place approximately every year, though a two-year gap occurred between the third and fourth timepoints due to COVID-19 restrictions. Demographic details of the participants at each timepoint are reported in Table 1.
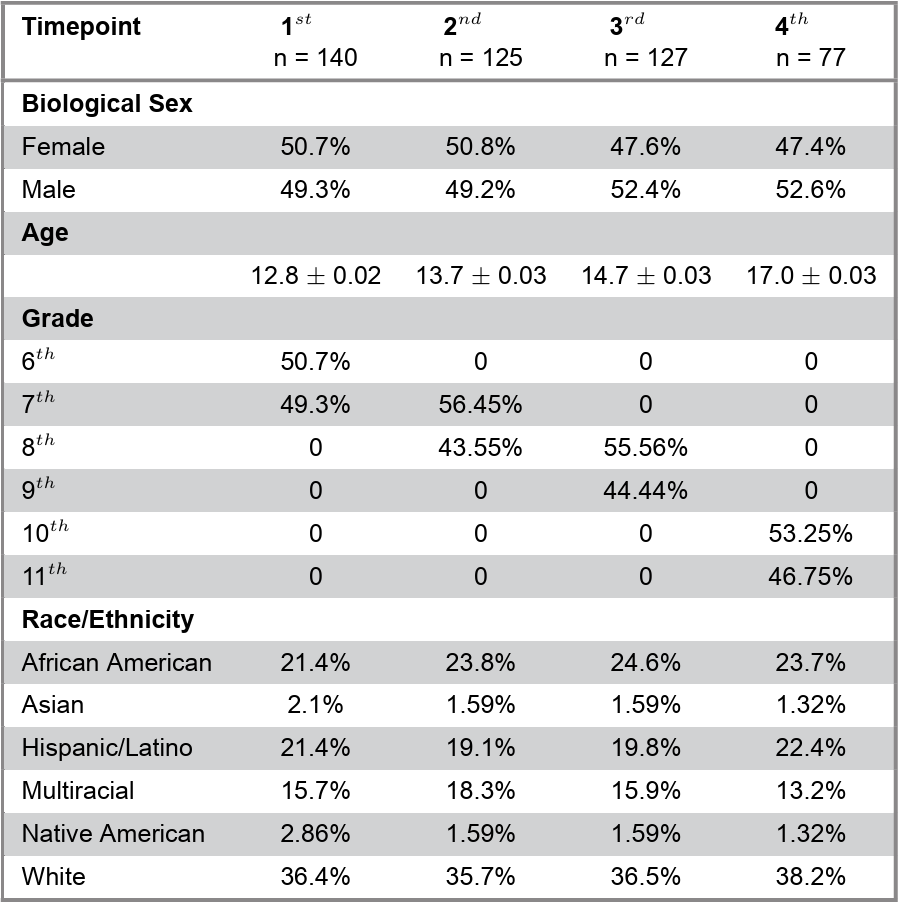
Table 1. Demographic information about the sample by timepoint.
Procedures and Measures
All study procedures were approved by the University of North Carolina at Chapel Hill’s Institutional Review Board. At each wave, participants provided informed assent/consent, demographic information, psychiatric symptom reports and completed tasks during a ~90-minute fMRI session. Depressive symptoms were measured using the Short Mood and Feelings Questionnaire (SMFQ; Angold et al. 1995; Eyre et al. 2021), a well-validated 13-item measure for pediatric and adolescent populations (Table 2). Participants rated how well each item reflected experiences during the previous two weeks (Not True [0], Sometimes True [1], Very True [2]). Responses were summed to yield a total score, with higher scores reflecting greater depressive symptom burden. Participants were compensated for their time and received additional monetary rewards based on task performance.

Table 2. SMFQ items.
MRI Data Acquisition
Scanning was conducted on a 3 Tesla Siemens Prisma MRI system. During each session, participants completed two functional runs of a Social Incentive Delay (SID) task and one functional run of a Cups risky decision-making task. Each run used a T2*-weighted, single-shot, gradient-echo, echo-planar imaging (EPI) sequence optimized for blood oxygenation level-dependent (BOLD) contrast (37 slices, 3mm thickness; voxel size 2.5 x 2.5 x 3mm; repetition time [TR] = 2000ms; echo time [TE] = 25ms; flip angle [FA] = 90˚; FOV = 230mm; 92 x 92 matrix). Each SID run comprised 195 volumes, and the Cups task run comprised 145 volumes. Slices were collected in the oblique axial plane that enhanced the coverage and reduced noise. T1-weighted structural images were obtained at each session with a sagittal T1-weighted magnetization-prepared rapid gradient-echo (MPRAGE) sequence (TR = 2400ms; TE = 2.22ms; 208 slices; voxel size = 0.8mm³).
Estimate of Dopamine-Related Neurophysiology
Functional data were preprocessed using the fMRI prep toolbox, including susceptibility distortion correction, skull-stripping, slice-time correction, registration to structural space and nonlinear warping to MNI (Montreal Neurological Institute) space. Following prior approaches, normalized T2*-weighted signal (nT2*w) was used as a proxy measure of tissue iron (Connor et al. 2009; Larsen et al. 2020b). For each timepoint, nT2*w was aggregated across three minimally preprocessed runs: two SID runs (~390 TRs) and one Cups run (~145 TRs). Whole-brain nT2*w maps were generated using the lncdtools toolbox (Jellen et al. 2013). TRs with greater than 0.3mm framewise displacement were censored, consistent with prior work (Beard 2003; Connor et al. 2009). On average, 6.7 ± 8.3% of all frames were excluded, with censoring rates unrelated to participant age (p = .166). Remaining volumes were z-score normalized using the fMRI prep-generated run-specific coverage map (Larsen et al. 2020a; Parr et al. 2022). Volumes were then aggregated across runs (~1224 volumes total) using voxel-wise medians that minimized the outlier effects, producing a single time-invariant nT2*w map for each participant at each wave. Regional averages were extracted from basal ganglia regions of interest (ROIs: caudate, nucleus accumbens, putamen and pallidum; Figure 3B), as defined by the Harvard-Oxford subcortical atlas, RRID:SCR_001476 (Anthony and Petronis 1995). Because nT2*w signal decreases with greater tissue iron, lower nT2*w values were indicated as a higher iron concentration and, in turn, greater dopaminergic activity (Wagner and Anthony 2002).
Analysis
Multilevel modeling (MLM) in the lmerTest R package was used to assess relationships between tissue iron and SMFQ scores across age, given its ability to account for multiple observations nested within individuals and both within- and between-person variability. Normality of SMFQ and tissue iron measurements by wave were assessed by obtaining skewness values with the e1071 R package and analyzing the normality of residuals to ensure conditions were met for MLM. Differential age-related changes in nT2*w values across the four basal ganglia ROIs were first assessed, controlled for mean framewise displacement and biological sex main and interaction effects (Model 1). Age and framewise displacement variables were grand-mean centered. All statistical tests were two-tailed. Significance was determined using a threshold of p < 0.05. Given significant ROI x age x sex interactions on nT2*w values, follow-up analyses of age x sex effects for each ROI separately were performed (Model 2). Then, to assess relationships between SMFQ scores and tissue iron across development, SMFQ scores were centered to each participant’s mean such that intra-individual changes in SMFQ, relative to one’s own mean, could be assessed while controlling between-person differences in average SMFQ score as well as mean framewise displacement and biological sex (Model 3).
Model 1. nT2w ~ Age_grandmc * ROI * Sex * meanFD_grandmc + (Age_grandmc | Subject) + (ROI | Subject) + (meanFD_grandmc | Subject)
Model 2. Region_nT2*w ~ Age_grandmc * Sex + meanFD_grandmc + (Age_grandmc | Subject) + (meanFD_grandmc | Subject)
Model 3. Region_nT2*w ~ Age_grandmc * SMFQ_submc + Age_grandmc * SMFQ_submean + meanFD_grandmc + Sex + (Age_grandmc | Subject) + (SMFQ_submc | Subject) + (meanFD_grandmc | Subject)
Post hoc
After completing the primary analyses and observing the complexity of within-person changes in tissue iron and depressive symptoms. Additional exploratory analyses were conducted for further understanding of the effects of depressive symptom severity during adolescence when disregarding changes in depressive symptoms over time. Specifically, participants’ average and maximum SMFQ results were used as predictors of variation in tissue iron across the observed developmental window. The relationships between average and maximum SMFQ scores and tissue iron were observed using Models 4 and 5 respectively. For post hoc analyses across the four basal ganglia ROIs, Bonferroni correction was applied to control for multiple comparisons (corrected p < 0.0125).
Model 4. Region_nT2*w ~ Age_grandmc * SMFQ_submean + Sex + meanFD_grandmc + (Age_grandmc | Subject) + (meanFD_grandmc | Subject)
Model 5. Region_nT2*w ~ Age_grandmc * SMFQ_submax + Sex + meanFD_grandmc + (Age_grandmc | Subject) + (meanFD_grandmc | Subject)
Results
When assessing tissue iron values across all timepoints and participants, distribution across the basal ganglia largely corresponded with findings from prior work. Specifically, the pallidum showed high tissue iron, while the nucleus accumbens displayed a wider range of nT2*w values (Larsen et al. 2020b; Parr et al. 2022; Figure 2). An omnibus test assessed significant interactions between age, sex and tissue iron (Model 1). This three-way interaction was significant, prompting follow-up analyses by ROI. A significant age x sex interaction was found in the caudate (0.003 ± 0.0; t[306.6] = 2.1; p = .037). Further tests revealed that age had a significant effect for females (-0.004 ± 0.0; t[150.0] = -3.3; p = .001), but not for males (p = .990; Figure 3a). Interactions in other regions were not significant: pallidum (p = .226), nucleus accumbens (p = .590), putamen (p = .067). Sex-related differences in age effects on tissue iron were specific to the caudate, with significant age-related increases observed exclusively in females.
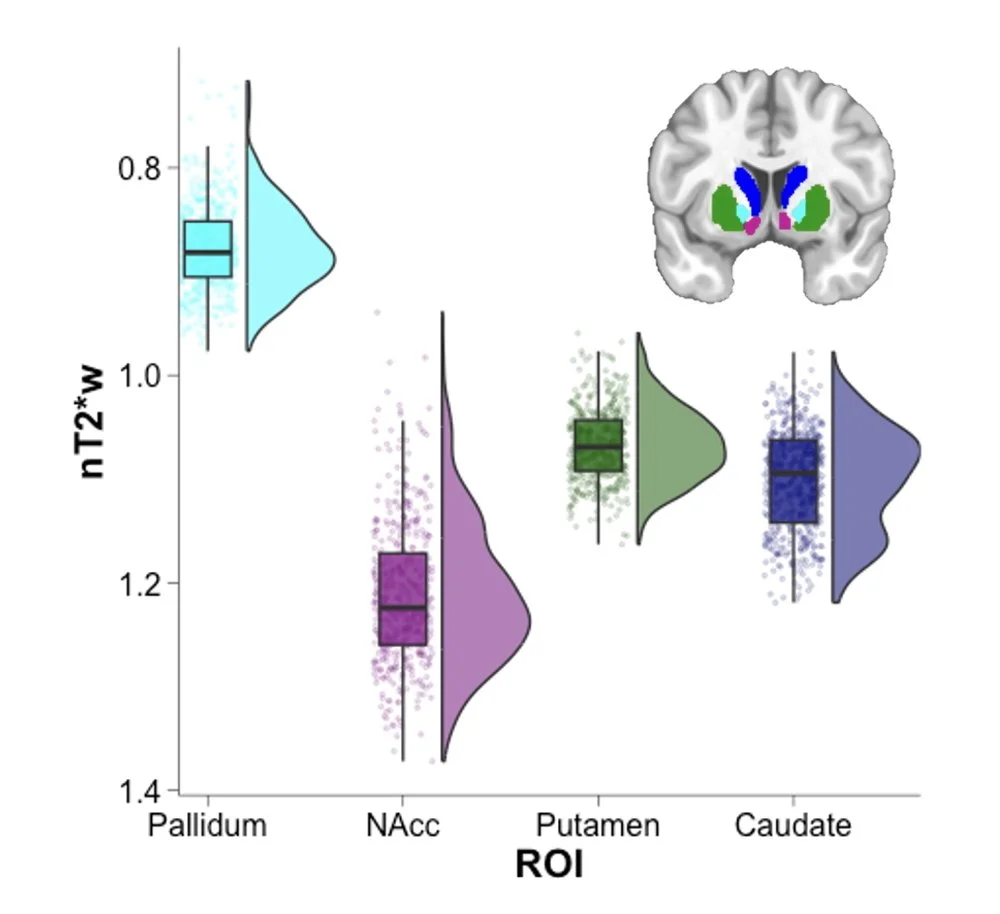
Figure 2. A boxplot showing the distribution of tissue iron in each ROI across participants. The graph has a reversed y-axis to reflect the inverse relationship between nT2*w and tissue iron.
When controlling for sex, there was a significant main effect of age in the pallidum, indicating that tissue iron increased with age (-0.005 ± 0.0; t[112.7] = -4.445; p = 1.99e-05; Figure 3b). Age-related increases were also significant in the nucleus accumbens (p = .003), putamen (p = 4.05e-12) and caudate (p = 8.45e-11).
The interaction between depressive symptoms (SMFQ) and age on overall basal ganglia nT2*w was not significant, whether using subject mean (p = .824) or mean-centered scores (p = .659). ROI-specific analyses also showed no significant effects (all p-values > .05).
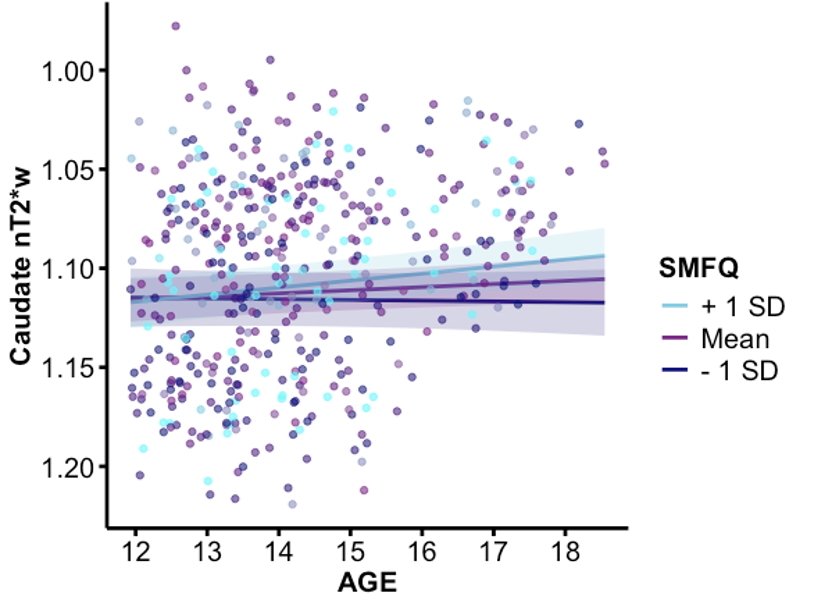
Figure 4. A plot of the interactions between age and caudate nT2*w by high, medium, and low SMFQ subject averages, controlling for biological sex and mean framewise displacement. The graph has a reversed y-axis to reflect the inverse relationship between nT2*w and tissue iron.
Post hoc
As the initial hypothesis was not supported, post hoc exploratory analyses of participants’ average and maximum depressive SMFQ scores were conducted to further understand the effect of depressive symptom severity on tissue iron development (Table 3). A significant age x average SMFQ score interaction was found in the caudate (p = .0111; Figure 4). This effect remained significant after Bonferroni’s correction for four ROIs (corrected p = .0444). No significant effects were observed in other basal ganglia regions, nor for age x maximum SMFQ interactions. These exploratory results suggest that greater average depressive symptoms may be associated with altered age-related changes in caudate tissue iron.
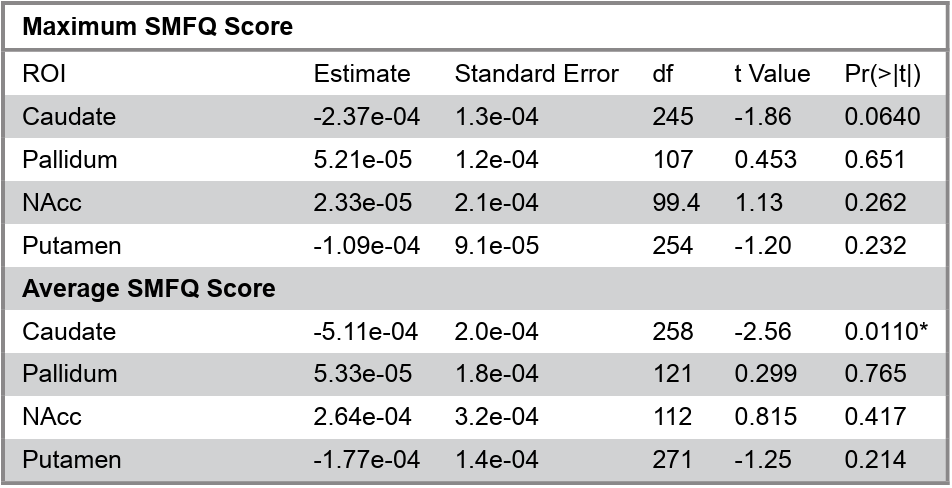
Table 3. Summary of the interactions between participants’ average and maximum SMFQ scores with age and nT2*w.
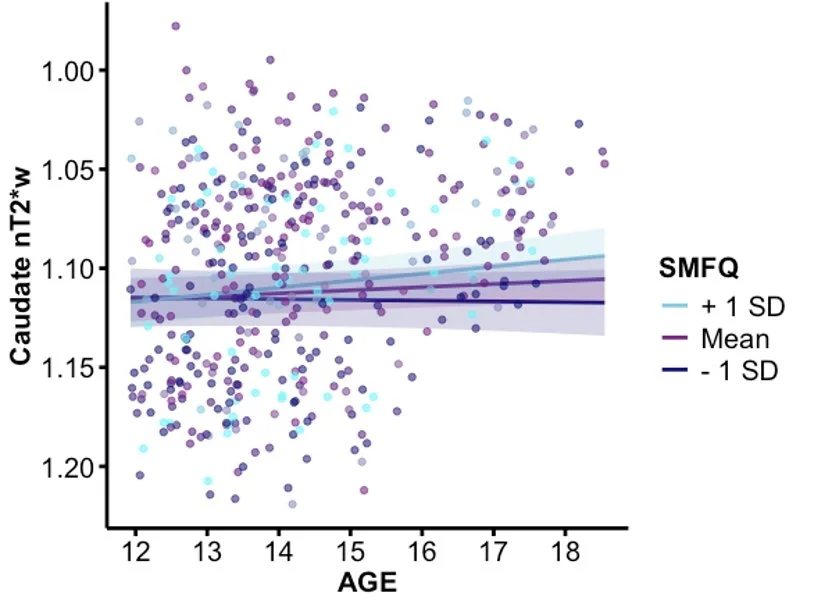
Figure 4. A plot of the interactions between age and caudate nT2*w by high, medium, and low SMFQ subject averages, controlling for biological sex and mean framewise displacement. The graph has a reversed y-axis to reflect the inverse relationship between nT2*w and tissue iron.
Discussion
This study examined how depressive symptoms relate to basal ganglia tissue iron development in adolescents. MLM did not reveal a significant interaction between tissue iron and depressive symptoms across development. Exploratory analyses, however, identified a significant interaction between age and averaged value in SMFQ scores in the caudate and a significant association between age and caudate tissue iron among females. These findings suggest individual differences in caudate tissue iron may be related to the depressive symptom severity, although the exact mechanisms remain unclear.
While our primary hypothesis was not supported, the exploratory finding in the caudate aligns with prior work showing that the caudate and other striatal regions play a key role in motivation and reward processing. Functional MRI studies have demonstrated altered BOLD activation in the caudate during reward anticipation among individuals with greater depressive symptoms, typically reflecting reduced responsiveness to rewarding cues (Pizzagalli et al. 2009). Established findings of blunted caudate BOLD signaling in depressed individuals, in combination with our results, indicate a potential relationship between dopamine signaling and depressive severity. As this study did not include BOLD measurement, however, further work is necessary to clarify this relationship.
The finding that higher depressive symptoms corresponded with higher tissue iron levels may reflect accelerated neural maturation, a phenomenon commonly observed following early life stress, which may have additional impacts on depression severity (Callaghan and Tottenham 2018; Fareri and Tottenham 2016; Smith and Pollak 2020; Saleh et al. 2017). Specifically, children who experienced early life stress exhibited decreased striatal activation during reward learning and increased anhedonia-like symptoms (Smith and Pollak 2020). This is not a conclusive result, however, given early life stress was not assessed in this study. Future research should investigate how interactive and additive effects of environmental influences and early life experiences impact tissue iron levels.
This study focused solely on depression and did not consider potential comorbid conditions, such as generalized anxiety disorder and attention deficit hyperactivity disorder, which are both linked to lower levels of striatal tissue iron (Shvarzman et al. 2022; Wu et al. 2023). Within-person differences, including sleep habits, self-esteem and iron deficiency, which may contribute to the severity of depression, were also not observed in this study (Kouros et al. 2016; Wu et al. 2023). As a result, the observed relationship between tissue iron levels and depressive symptoms may be better explained by these factors. Future studies should seek to control for these variables, either in experimental design or as a part of statistical analyses, to isolate the relationship between depressive symptoms and adolescent tissue iron development.
It is important to note that analyses of average and maximum SMFQ scores address a fundamentally different question to that of the original hypothesis. This study initially sought to examine developmental changes in depressive severity, but significance emerged only when exploring general differences between individuals across timepoints. Examining tissue iron trajectories and average depressive severity may be more important, however, as the rank-order stability of depression increases throughout adolescence and conclusions with this data could have greater implications beyond this window (Prenoveau et al. 2011; Son et al. 2023). Grouping within subjects in these analyses may have also contributed to greater power, thus increasing the significance of the results. Future work should explore if these findings are generalizable to clinical samples.
Our identification of a significant relationship between age and caudate tissue iron in females replicates past studies’ findings related to sex differences in brain development. One study found adult females had significantly lower iron across several regions (Bartzokis et al. 2007). It is theorized that estrogen and higher rates of iron deficiency in females may be related to sex differences in tissue iron and mood disorders, as women are twice as likely to be diagnosed with depression than men (Balzer et al. 2015; Eid et al. 2019; Grubić Kezele and Ćurko-Cofek 2020; Larsen et al. 2020a;). Poon et al. (2019) observed an inverse relationship between estradiol and NAcc-prefrontal region connectivity and another inverse relationship between testosterone and dorsolateral prefrontal cortex activation. As regional connectivity in these areas is closely tied to reward functioning, pubertal hormones may have a measurable effect on depressive severity via anhedonia-related symptoms (Gupta et al. 2024). Additional research is necessary to determine if there is a direct relationship between these hormones and depression or if a mediating factor, like tissue iron, is involved.
The potential involvement of estrogen in depression and tissue iron development and the natural changes in estrogen levels across puberty, coupled with the general lack of research on female and adolescent participants, make future research on this developmental window all the more necessary (Balzer et al. 2015; Poon et al. 2019). Specifically, future studies should observe differential relationships with tissue iron and depression separately for female and male adolescents with a larger sample size. Observing factors like sex hormones, menstruation-related iron loss and pubertal timing in relation to tissue iron and depressive symptom development will significantly enhance understanding of the impact of biological factors on psychopathology risk and inform future biomedical interventions for disorders like depression.
The findings of this study are fundamentally limited by the fact that all measures of dopaminergic activity were collected indirectly. While tissue iron has demonstrated associations with dopaminergic development in prior literature, it is, nonetheless, an indirect, correlational measure (Larsen et al. 2020b). The development of less invasive methods of direct dopamine assessment for adolescents would strengthen these findings, improve the validity of future work and may reveal different relationships between dopaminergic activity, development and psychopathology.
Conclusion
Exploratory analyses revealed a relationship between depressive symptoms and caudate tissue iron, particularly for females. These results suggest that individual differences in caudate tissue iron may be an indicator for adolescent depressive severity, though further work is needed to support this association. The lack of relationships found between within-person changes in depressive symptoms and tissue iron suggests that more stable factors, such as early life stress, hormonal levels and peripheral iron levels, may play a significant role in the progression of depressive symptoms.
This study provides a valuable framework for future work, which can elucidate the impacts of sex and external factors on tissue iron and depression. Additional research is necessary to confirm the validity of tissue iron as an indicator of dopaminergic function and development, identify critical influences on adolescent mental health and inform effective, targeted interventions to mitigate the long-term effects of depression.
Acknowledgements
First, I want to thank Dr. Jessica Flannery for her support as my advisor throughout this project. Her expertise, patience and advice were central to the success of this work, and I am so thankful to have had the opportunity to work with her. I would also like to thank Dr. Eva Telzer and the entire Developmental Social Neuroscience Lab at UNC-Chapel Hill, which supported my thesis and provided the data necessary for my analyses. Further, I would like to thank Dr. Telzer and Dr. Annie Maheux for reading my thesis and participating in my defense board. I am also incredibly grateful to Dr. Adam Goldstein for his continuing research mentorship and support throughout preparing this manuscript for publication.
Conflicts of Interest/Disclosure
The author of this manuscript has no conflicts of interest to disclose.
References
Angold, A., Costello, E.J., Messer, S.C. and Pickles, A. (1995) ‘Development of a short questionnaire for use in epidemiological studies of depression in children and adolescents’, International Journal of Methods in Psychiatric Research, 5(4), 237-249.
Anthony, J.C. and Petronis, K.R. (1995) ‘Early-onset drug use and risk of later drug problems’, Drug and Alcohol Dependence, 40(1), 9-15.
Balzer, B.W.R., Duke, S.A., Hawke, C.I. and Steinbeck, K.S. (2015) ‘The effects of estradiol on mood and behavior in human female adolescents’, European Journal of Pediatrics, 174(3), 289-298.
Bartzokis, G., Tishler, T.A., Lu, P.H., Villablanca, P., Altshuler, L.L., Carter, M., Huang, D., Edwards, N. and Mintz, J. (2007) ‘Brain ferritin iron may influence age- and gender-related risks of neurodegeneration’, Neurobiology of Aging, 28(3), 414-423.
Beard, J. (2003) ‘Iron Deficiency Alters Brain Development and Functioning’, The Journal of Nutrition, 133(5), 1468S-1472S.
Callaghan, B.L. and Tottenham, N. (2018) ‘The stress Acceleration Hypothesis: effects of early-life adversity on emotion circuits and behavior’, Current Opinion in Behavioral Sciences, 7, 76-81.
Connor, J.R., Wang, X.S., Allen, R.P., Beard, J.L., Wiesinger, J.A., Felt, B.T. and Earley, C.J. (2009) ‘Altered dopaminergic profile in the putamen and substantia nigra in restless leg syndrome’, Brain: A Journal of Neurology, 132(Pt 9), 2403-2412.
Cooper, J.A., Arulpragasam, A.R. and Treadway, M.T. (2018) ‘Anhedonia in depression: biological mechanisms and computational models’, Current Opinion in Behavioral Sciences, 22, 128-135.
Daubner, S.C., Le, T. and Wang, S. (2011) ‘Tyrosine hydroxylase and regulation of dopamine synthesis’, Archives of Biochemistry and Biophysics, 508(1), 1-12.
Davey, C.G., Yücel M. and Allen, N.B. (2008) ‘The emergence of depression in adolescence: development of the prefrontal cortex and the representation of reward’, Neuroscience and Biobehavioral Reviews, 32(1), 1-19.
Eid, R.S., Gobinath, A.R. and Galea, L.A.M. (2019) ‘Sex differences in depression: Insights from clinical and preclinical studies’, Progress in Neurobiology, 176, 86-102.
Eyre, O., Bevan-Jones, R., Agha, S.S., Wootton, R.E., Thapar, A.K., Stergiakouli, E., Langley, K., Collishaw, S., Thapar, A. and Riglin, L. (2021) ‘Validation of the short Mood and Feelings Questionnaire in young adulthood’, Journal of Affective Disorders, 294, 883-888.
Fareri, D.S. and Tottenham, N. (2016) ‘Effects of early life stress on amygdala and striatal development’, Developmental Cognitive Neuroscience, 19, 233-247.
Flannery, J.S., Parr, A.C., Lindquist, K.A. and Telzer, E.H. (2025) ‘Developmental changes in dopamine-related neurophysiology and associations with adolescent substance use and incentive-boosted cognitive control’, Developmental Cognitive Neuroscience, 85, 101594.
Gabbay, V., Ely, B.A., Li, Q., Bangaru, S.D., Panzer, A.M., Alonso, C.M., Castellanos, F.X. and Milham, M.P. (2013) ‘Striatum-based circuitry of adolescent depression and anhedonia’, Journal of the American Academy of Child and Adolescent Psychiatry, 52(6), 628-41.e13.
Galvan, A. (2010) ‘Adolescent development of the reward system’, Frontiers in Human Neuroscience, 4, 6.
Grubić Kezele, T. and Ćurko-Cofek, B. (2020) ‘Age-Related Changes and Sex-Related Differences in Brain Iron Metabolism’, Nutrients, 12(9), 2601.
Gupta, T., Eckstrand, K.L. and Forbes, E.E. (2024) ‘Annual Research Review: Puberty and the development of anhedonia – considering childhood adversity and inflammation’, Journal of Child Psychology and Psychiatry, and Allied Disciplines, 65(4), 459-480.
Hill, J.M. and Switzer, R.C. (1984) ‘The regional distribution and cellular localization of iron in the rat brain’, Neuroscience, 11(3), 595-603.
Jellen, L.C., Lu, L., Wang, X., Unger, E.L., Earley, C.J., Allen, R.P., Williams, R.W. and Jones, B.C. (2013) ‘Iron deficiency alters expression of dopamine-related genes in the ventral midbrain in mice’, Neuroscience, 252, 13-23.
Kouros, C.D., Morris, M.C. and Garber, J. (2016) ‘Within-Person Changes in Individual Symptoms of Depression Predict Subsequent Depressive Episodes in Adolescents: A Prospective Study’, Journal of Abnormal Child Psychology, 44(3), 483-494.
Larsen, B., Bourque, J., Moore, T.M., Adebimpe, A., Calkins, M.E., Elliott, M.A., Gur, R.C., Gur, R.E., Moberg, P.J., Roalf, D.R., Ruparel, K., Turetsky, B.I., Vandekar, S.N., Wolf, D.H., Shinohara, R.T. and Satterthwaite, T.D. (2020a) ‘Longitudinal development of brain iron is linked to cognition in youth’, The Journal of Neuroscience, 40(9), 1810-1818.
Larsen, B., Olafsson, V., Calabro, F., Laymon, C., Tervo-Clemmens, B., Campbell, E., Minhas, D., Montez, D., Price, J. and Luna, B. (2020b) ‘Maturation of the human striatal dopamine system revealed by PET and quantitative MRI’, Nature Communications, 11(1), 846.
Laviola, G., Macrì, S., Morley-Fletcher, S. and Adriani, W. (2003) ‘Risk-taking behavior in adolescent mice: psychobiological determinants and early epigenetic influence’, Neuroscience and Biobehavioral Reviews, 27(1-2), 19-31.
Luking, K.R., Pagliaccio, D., Luby, J.L. and Barch, D.M. (2016) ‘Reward processing and risk for depression across development’, Trends in Cognitive Sciences, 20(6), 456-468.
Mills, K.L., Siegmund, K.D., Tamnes, C.K., Ferschmann, L., Wierenga, L.M., Bos, M.G.N., Luna, B., Li, C. and Herting, M.M. (2021) ‘Inter-individual variability in structural brain development from late childhood to young adulthood’, Neuroimage, 242, 118450.
Modrzejewska, R., Bomba, J., Cofór, P. and Pac, A. (2022) ‘Depressive symptoms in adolescence and quality of life 17 years later – follow-up study’, Psychiatria Polska, 56(1), 51-61.
Parr, A.C., Calabro, F., Tervo-Clemmens, B., Larsen, B., Foran, W. and Luna, B. (2022) ‘Contributions of dopamine-related basal ganglia neurophysiology to the developmental effects of incentives on inhibitory control’, Developmental Cognitive Neuroscience, 54, 101100.
Pizzagalli, D.A., Holmes, A.J., Dillon, D.G., Goetz, E.L., Birk, J.L., Bogdan, R., Dougherty, D.D., Iosifescu, D.V., Rauch, S.L. and Fava, M. (2009) ‘Reduced caudate and nucleus accumbens response to rewards in unmedicated individuals with major depressive disorder’, The American Journal of Psychiatry, 166(6), 702-710.
Poon, J.A., Niehaus, C.E., Thompson, J.C. and Chaplin, T.M. (2019) ‘Adolescents’ pubertal development: Links between testosterone, estradiol, and neural reward processing’, Hormones and Behavior, 114, 104504.
Prenoveau, J.M., Craske, M.G., Zinbarg, R.E., Mineka, S., Rose, R.D. and Griffith, J.W. (2011) ‘Are anxiety and depression just as stable as personality during late adolescence? Results from a three-year longitudinal latent variable study’, Journal of Abnormal Psychology, 120(4), 832-843.
Rappaport, B.I., Kandala, S., Luby, J.L. and Barch, D.M. (2020) ‘Brain reward system dysfunction in adolescence: current, cumulative, and developmental periods of depression’, The American Journal of Psychiatry, 177(8), 754-763.
Saleh, A., Potter, G.G., McQuoid, D.R., Boyd, B., Turner, R., MacFall, J.R. and Taylor, W.D. (2017) ‘Effects of early life stress on depression, cognitive performance and brain morphology’, Psychological Medicine, 47(1), 171-181.
Shah, H.E., Bhawnani, N., Ethirajulu, A., Alkasabera, A., Onyali, C.B., Anim-Koranteng, C. and Mostafa, J.A. (2021) ‘Iron Deficiency-Induced Changes in the Hippocampus, Corpus Striatrum, and Monoamines Levels That Lead to Anxiety, Depression, Sleep Disorders, and Psychotic Disorders’, Cureus, 13(9), e18138.
Shorey, S., Ng, E.D. and Wong, C.H.J. (2022) ‘Global prevalence of depression and elevated depression symptoms among adolescents: A systematic review and meta-analysis’, The British Journal of Clinical Psychology, 61(2), 287-305.
Shvarzman, R., Crocetti, D., Rosch, K.S., Li, X. and Mostofsky, S.H. (2022) ‘Reduced basal ganglia tissue-iron concentration in school-age children with attention-deficit/hyperactivity disorder is localized to limbic circuitry’, Experimental Brain Research, 240(12), 3271-3288.
Smith, K.E. and Pollak, S.D. (2020) ‘Early life stress and development: potential mechanisms for adverse outcomes’, Journal of Neurodevelopmental Disorders, 12(1), 34.
Son, S., Lee, H. and Jang, Y. (2023) ‘Continuity and Stability of Child and Adolescent Depressive Symptoms in South Korea: A Meta-analysis of Longitudinal Studies’, Journal of Youth and Adolescence, 52(3), 598-618.
Stewart, R. and Hirani, V. (2012) ‘Relationship between depressive symptoms, anemia, and iron status in older residents from a national survey population’, Psychosomatic Medicine, 74(2), 208-213.
Urošević S., Collins, P., Muetzel, R., Lim, K. and Luciana, M. (2012) ‘Longitudinal changes in behavioral approach system sensitivity and brain structures involved in reward processing during adolescence’, Developmental Psychology, 48(5), 1488-1500.
Vahdat Shariatpanaahi, M., Vahdat Shariatpanaahi, Z., Moshtaaghi, M., Shahbaazi, S.H. and Abadi, A. (2007) ‘The relationship between depression and serum ferritin level’, European Journal of Clinical Nutrition, 61(4), 532-535.
Wagner, F.A. and Anthony, J.C. (2002) ‘From first drug use to drug dependence; developmental periods of risk for dependence upon marijuana, cocaine, and alcohol’, Neuropsychopharmacology, 26(4), 479-488.
Wahlstrom, D., Collins, P., White, T. and Luciana, M. (2010) ‘Developmental changes in dopamine neurotransmission in adolescence: behavioral implications and issues in assessment’, Brain and Cognition, 72(1), 146-159.
Wierenga, L., Langen, M., Ambrosino, S., van Dijk, S., Oranje, B. and Durston, S. (2014) ‘Typical development of basal ganglia, hippocampus, amygdala and cerebellum from age 7 to 24’, Neuroimage, 96, 67-72.
Wu, Q., Ren, Q., Meng, J., Gao, W.J. and Chang, Y.Z. (2023) ‘Brain iron homeostasis and mental disorders’, Antioxidants, 12(11), 1997.
Yao, S., Zhong, Y., Xu, Y., Qin, J., Zhang, N., Zhu, X. and Li, Y. (2017) ‘Quantitative Susceptibility Mapping Reveals an Association between Brain Iron Load and Depression Severity’, Frontiers in Human Neuroscience, 11, 442.
Yi, S., Nanri, A., Poudel-Tandukar, K., Nonaka, D., Matsushita, Y., Hori, A., Mizoue, T. (2011) ‘Association between serum ferritin concentrations and depressive symptoms in Japanese municipal employees’, Psychiatry Research, 189(3), 368-372.
Yoshida, J., Oñate, M., Khatami, L., Vera, J., Nadim, F. and Khodakhah, K. (2022) ‘Cerebellar contributions to the basal ganglia influence motor coordination, reward processing, and movement vigor’, The Journal of Neuroscience, 42(45), 8406-8415.