

Authors: Uma Goyal (1), Laurie Bryant (2)
Institution: University of Arizona (1), Southwest Baptist University (2)
Date: May 2007
Abstract
Body size (based on weaning weights) in mammals has shown varying importance for their reproductive and survival success. Therefore, size can be influential among an individual's interactions with the environment. This field study is the first to compare the body size of prairie voles (Microtus ochrogaster) and its effect on their interaction with others and overall survival while controlling the density of animals in each enclosure. We hypothesized that a larger body size for prairie voles promotes opposite-sex interactions, pregnancy, and survival in higher densities. However, size does not affect these indexes in moderate-density enclosures. To determine if there was an association between these variables, lab reared prairie voles were weighed at weaning (21 days old) and then released into high and moderate density field enclosures. We followed the animals using a grid and nest trapping system, and then used statistical tests in order to determine the prairie voles' actions. We found that body size had no significant correlation to the prairie voles' behavior in these categories. These results show that there may be other causes to the difference between voles' activities. Understanding the factors that influence an animal's reproductive and survival success will allow us to see how they interact with other animals and their environment as a whole, which can help us to better understand other species.
Introduction
In small mammals, particularly rodents, body size seems to play a pivotal role in an individual male or female's reproductive success. Larger body size may benefit males and females due to risks of predation. In one study, larger field voles dispersed further than smaller voles in order to avoid carnivores (Norrdahl and Korpimaki, 1998). In another study, a lower impact of predation and higher survival rates were recorded for many larger alpine bank voles. Also, a larger body size for males resulted in increased sexual selection due to increased survival (Yoccoz and Mesnager, 1998). Larger female prairie voles may have superior energy stores than smaller voles, which would be beneficial during lactation when energy intake is significantly higher than that of non-reproductive voles (He-Liu et al., 2003). In addition, larger female prairie voles may be better able to protect their offspring since maternal aggression is an important part of vole survival (Bowler et al., 2002).
However, there were also studies that concluded that there is no clear advantage of body size. In yellow-pine chipmunks, male reproductive success was independent of body size and was attributed to other behavioral components such as dominance and running speed (Schulte-Hostedde et al., 2002). It has also been reported that body size is dependent on age and season. Therefore, many small voles may not have had enough time to become large (Boonstra and Krebs, 1979).
Of previous studies examined, the effects of adult body size on various factors linked to reproductive success, (Solomon, 1993) found that adult body size in prairie voles was correlated with body mass at weaning. Females that were larger at weaning had offspring that grew faster prior to weaning (Solomon, 1994). Increased growth rates may allow offspring to become independent sooner. In addition, offspring that were larger at weaning had higher survival rates immediately after weaning than offspring with lower body mass at weaning (Solomon, 1991). Finally, in the laboratory, prairie voles of larger body size had a better chance of being preferred as a partner than prairie voles of smaller size (Solomon, 1993).
Yet this same observation seen in laboratory prairie voles is not confirmed by descriptive field data (Getz et al., 2000). Getz et al. (2000) found that there was no correlation between body size and reproductive success. Prairie voles are believed to be socially monogamous, and thus the role of body size with mate choice and standard of living was studied in a field setting in order to determine their behavior under natural conditions. The purpose of this research is to determine whether weaning weight has an effect on reproduction and survival. Since results of previous studies on the effects of body size in rodents are not consistent, prairie voles (Microtus ochragaster) were studied by grid and nest trapping in order to determine any possible relationship within a field setting. In addition, we examined the effect that weaning weight has at high and moderate population densities. To determine if there was an association between body size, reproduction and survival, lab reared prairie voles were weighed at weaning (21 days old) and released into field enclosures. We tracked the prairie voles using a grid and nest trapping system, and then used statistical tests in order to determine their behaviors. We found that body size had no significant correlation to the prairie voles' actions in these indexes, which indicates that there may be other causes to the difference between voles' activities. Thus, the results from this study allow us to better understand the interactions of these animals within their natural environment.
Materials and Methods
This experiment was conducted at Miami University's Ecology Research Center near Oxford, Ohio (39°30'N, 88°44'W). We collected data from June through August 2005. Prairie vole populations were established in ten 0.1-ha experimental enclosures. Enclosures were made of 20-gauge galvanized steel reaching 75 cm above and 45 cm below ground to prevent movement of rodents among enclosures. The enclosures contain a mixture of bluegrass (Poa pratensis), clover (Trifolium spp.), fescue (Festuca sp.), timothy (Phleum sp.), and ryegrass (Elymus sp.) in order to provide cover and a food source. Vegetation was sampled in June 2005. A 1 m wide strip of vegetation was mowed periodically around the inside of the enclosures in order to discourage digging near the walls (Solomon and Jaquot, 2002). The enclosures were surrounded by an electric fence in order to prevent terrestrial predators like raccoons and feral cats from entering.
Prior to the current experiment, all voles were removed by live-trapping until there was a consistent 3 days of no captures prior to release of the founders. F2-generation laboratory reared voles from wild voles caught in east central Illinois were released into the enclosures. The wild voles were randomly paired to breed and the F1-generation was also paired at the age of 6 months to 2 years of age. The F2 offspring were weaned, uniquely marked, weighed to 0.1 g, and measured from nose to base of tail at 21 days of age. They were removed from their parents' cage and housed with siblings under the same conditions as their parents. The voles were housed under a 14L:10D lighting schedule in plastic 46x25x20-cm cages with recycled paper bedding (Cell Sorb Plus, Fangman Specialties, Cincinnati, Ohio). They were given rodent chow (Purina Mills, Richmond, Ind.) and water ad libium.
The F2-generation (founding voles) was assigned to enclosures in order to minimize the number of siblings in each enclosure, distribute ages, and maximize diversity. The founding voles were between the ages of 30 days and 4 months at the time of release. They were released in the middle of the enclosures at dusk and given a week to adapt to the new environment.
Enclosures were randomly assigned to high and moderate density treatments. Six high-density enclosures were created containing 12 females and 12 males in each. Four moderate-density enclosures were also created containing 4 females and 4 males in each. Every female was weighed each time she was trapped and every male was weighed each week to 0.1g using a Pesola spring balance. We used a combination of grid trapping and nest trapping to collect data. Grid trapping allowed us to track the density of voles, and nest trapping allowed us to determine which voles were living together. Grid trapping occurred for the first 3 weeks in order to determine population density and allow us to determine the location of females. Then nest trapping and grid trapping alternated each week for 9 weeks. Finally, weeks 10-12 were entirely nest trapping.
For grid trapping, trap stations were located in a grid with stakes 5.3 m apart in a 5x5 arrangement with 25 Ugglan multiple capture traps (Grabnab, Sweden). Grid traps were set at 2000 h on Sundays. For the next two days, traps were checked at 0700, set at 1800, then checked and reset at 2100. Wednesday the traps were checked at 0700. For nest trapping, 3 Ugglan traps were placed at each nest location. Traps were set 2000 h on Sundays, and checked at 0700 the next morning, set at 1800, then checked and reset at 2100 Monday through Friday. Friday night the traps were not reset. Radio tracking was used to track the adult females to their nests. Radio collars (Holohill Co., Fl.) weigh less than 3 g (less than 10% of the animal's body weight). Collars were put on while females were under anesthesia (Iso-fluorane, Abbott Lab, Germany). For each tracking day, collared females were located within their designated enclosures. If a female was located in a specific area and a nest was found, it would be marked with a white flag. The white flag was designated a grid coordinate number for future nest trapping. After a week, the collars were removed using scissors.
When an animal was caught, the unique ID, sex, reproductive status, and age were recorded. Females were weighed once during each capture day and males were weighed once each capture week. Newly captured voles were uniquely marked and weighed as described above.

article_1033_order_0
The proportional hazards survival regression analysis was conducted using the Cox Proportional Hazards Survival Regression program at StatPages.net.and all other statistical analyses were conducted using StatView 5.0.1(SAS Institute Inc., Cary, NC). Proportion hazards survival regression analysis (Lawless, 1982) was used to analyze male and female survival over the 15-week study. Mean weaning weight of voles that survived and died during the study were analyzed with a t-test. Simple linear regression was used to compare the number of females a male was trapped with and male weaning weight. A t-test or ANOVA was used to compare the weaning weight of females that reproduced at least once versus did not reproduce. All analyses were conducted for all individuals of each sex combined across the two densities as well as for each density separately. All statistical analyses will be one-tailed because of a priori predictions. Differences will be considered statistically significant if p ≤ 0.05.
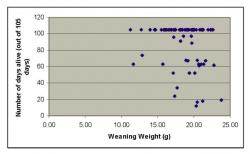
article_1033_order_1
Results
Survival
When considering all individuals from both densities, there was no significant relationship between weight at weaning and survival for males (χ2=1.9; P=0.17; figure 1) or females ((χ2=2.1; P=0.15; figure 2). There was also no significant relationship between weight at weaning and survival at high density (males: (χ2=1.0; P=0.33; females: (χ2=0.4; P=0.53) or moderate density (males: (χ2=1.3; P=0.25; females: (χ2=3.6; P=0.06). There was no significant difference in the mean weight at weaning of the voles that survived compared to those that died for either sex or density (Table 1). Thus, weight at weaning did not have a significant effect on survival in either male or female voles.

article_1033_order_2
Reproduction
There was no relationship between the number of different females a male was trapped with and weaning weight when considering males across the two densities (F1,80=1.99; P=0.16) or at high (F1,65=1.24; P=0.27) or moderate (F1,14=2.66; P=0.13) density. There was no significant difference in the weaning weight of females that reproduced at least once versus did not reproduce when considering all females or just females at high density (Table 2). All females reproduced at the moderate density. Likewise there was no significant difference in the weaning weight of females that did not reproduce, reproduced once and reproduced more than once (Table 3) when considering all females or the densities separately.
*P-values underpowered.

article_1033_order_3
Discussion
Lab reared prairie voles were weaned (weight at 21 days old) and released into field enclosures. The prairie voles were tracked using a grid and nest trapping system, and statistical tests were used in order to determine correlations between body size, reproduction and survival. We did not find any relationship between body sizes of prairie voles at weaning and survival or reproduction indexes. Thus, these data suggest that within a semi-natural field setting, neither male nor female prairie voles demonstrate reproductive or survival advantages based on weaning weights.

article_1033_order_4
This conclusion is not consistent with our hypothesis that larger body size increases survival and fitness in high-density enclosures, but the data does agree with our hypothesis based on moderate-density enclosures. This suggests that body size behavior at weaning is not a major determining factor in this socially monogamous mammal's mating behavior and survival in a more natural environment. It also suggests that the density of prairie voles does not significantly interact with weaning weight to affect survival or indexes of reproduction. Thus, weaning weight was not a major factor influencing survival or indexes of reproduction at moderate or high densities.
We had expected that large size at weaning would be important for males and females since weaning weight is highly correlated with adult weight of prairie voles in the lab (Solomon, 1993). We predicted that increased size would be beneficial for both sexes so that mates would be better able to defend territory and offspring. In the lab, prairie vole males and females preferred to associate with significantly larger partners (Solomon, 1993). Also, larger northern male grasshopper mice had increased testes size during the breeding season, while the monogamous males would have smaller testes as a result of infrequent copulations. Therefore, larger males mated with several different females (Goheen et al., 2003). In contrast, extra-large California vole males were found in areas that lacked females and poorer habitats. The Chitty Effect (extra-large body size in rodents causes high fitness under increasing and high densities) was not shown to occur in California voles, and they did not have enhanced fitness under conditions of rapid population growth and high densities. It was suggested that weight gain was seasonal and a result of density regulation, and extra-large voles sought habitats with tall and dense vegetation to avoid predators. A "Big Wimp Hypothesis" suggested that extra-large voles value predator avoidance more than reproductive or social dominance (Lidicker and Ostfeld, 1991). Thus, there were several variances on small mammal behaviors when associated with body size.
This research is important in determining the effects of weaning weight on the dispersal, survival, and reproductive success of socially monogamous small mammals within a field setting. Therefore, we can determine the indexes and behaviors (i.e., aggression, mate choice) that cause characteristics such as body size to vary so widely among small mammals in a natural environment.
Our results showed the lack of association between body size, density of animals within enclosures, and their actions that caused the rejection of our hypothesis and the suggestion that body size does not have a significant bearing on behavior. Future results that may support this suggestion would be similar field studies using other small mammals, having more extreme differences in weaning weights, or creating higher density differences between enclosures. Also, weaning weight may matter but the range of weights may need to be more extreme among enclosure founders in order to see significant behavioral effects.
Since data gathered from this experiment was collected by teams of individuals whose schedules rotated biweekly, this variance may have led to human error in field observations. In addition, our measurement of reproduction was somewhat indirect because basis of pregnancy was decided on physical characteristics such as mating plugs and extreme weight gain. Therefore, it would be better to also have genetic data on actual parenting of offspring in order to measure fitness. Also, a few founding voles died due to natural and accidental causes which altered our original sample size.
Conclusion:
Overall, the results from this experiment demonstrate that body size has no significant effect on reproduction and survival in a natural environment for small monogamous mammals. The next step to further prove this experimental result is to analyze effects of body size on other indexes such as wounding, aggression, and habitat vegetation quality in moderate and high-density enclosures. Thus, this research allows us to better understand the behavior of small mammal species and determine the effects of body size on their dispersal, survival, and reproduction success.
Acknowledgments
We would like to thank the following people for their work on this research project: Kristin Lucia, Ashley Richmond, Caryn Sorge, Michelle Edwards, Shannon Williams, and Tony Fries. Also, we would like to thank Miami University's NSF-REU program "Ecology in Human-Dominated Landscapes" and NSF DEB-0316818.
Works Cited
Boonstra, R and CJ Krebs (1979) Viability of large- and small-sized adults in fluctuating
vole populations. Ecology 60: 567-573.
Bowler, MB et al. (2002) Social factors regulate female-female aggression and
affiliation in prairie voles. Physiology and Behavior 76: 559-566.
Getz, LL et al. (2000) Nestling survival and population cycles in the prairie vole,
Microtus ochrogaster. Canadian Journal of Zoology 78 (10): 1723-1731.
Goheen, JR et al. (2003) Effect of body size on reproductive characteristics of the
northern grasshopper mouse in north-central Kansas. Southwestern Naturalist 48 (3): 427-431.
He-Liu et al. (2003) Energy requirements during reproduction in female brandt's voles
(Microtus brandii). Journal of Mammalogy 84(4): 1410-1416.
Lawless, JF (1982) Statistical Models and Methods for Lifetime Data. J. Wiley &
Sons, New York.
Lidicker, WZ Jr. and RS Ostfeld (1991) Extra-large body size in California voles
causes and fitness consequences. Oikos. 61(1): 108-121.
Norrdahl, K and E Korpimaki (1998) Does mobility or sex of voles affect risk of
predation by mammalian predators? Ecology (Washington DC) 79 (1): 226-232.
Schulte-Hostedde, AI et al. (2002) Female-biased sexual size dimorphism in the yellow-
pine chipmunk (Tamias amoenus): Sex-specific patterns of annual reproductive success and survival. Evolution 56 (12): 2519-2529.
Solomon, NG (1993) Body size and social preferences of male and female prairie
voles, Microtus ochrogaster. Animal Behaviour 45: 1031-1033.
Solomon, NG (1994) Effect of the pre-weaning environment on subsequent reproduction
in prairie voles, Microtus ochrogaster. Animal Behaviour 48: 331.
Solomon, NG (1991) Indirect fitness benefits associated with philopatry in juvenile
prairie voles. Behavioral Ecology and Sociobiology 29: 277-282.
Solomon, NG and J Joseph (2002) Characteristics of resident and wandering prairie
voles, Microtus ochrogaster. Canadian Journal of Zoology 80 (5): 951-955.
Yoccoz, NG and S Mesnager (1998) Are alpine bank voles larger and more sexually
dimorphic because adult voles survive better? Oikos 82 (1): 85-98.